First Report on the Acrobat Ant Crematogaster scutellaris Storing Live Aphids in Its Oak-Gall Nests

 , ,
, ,  ,
,  , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
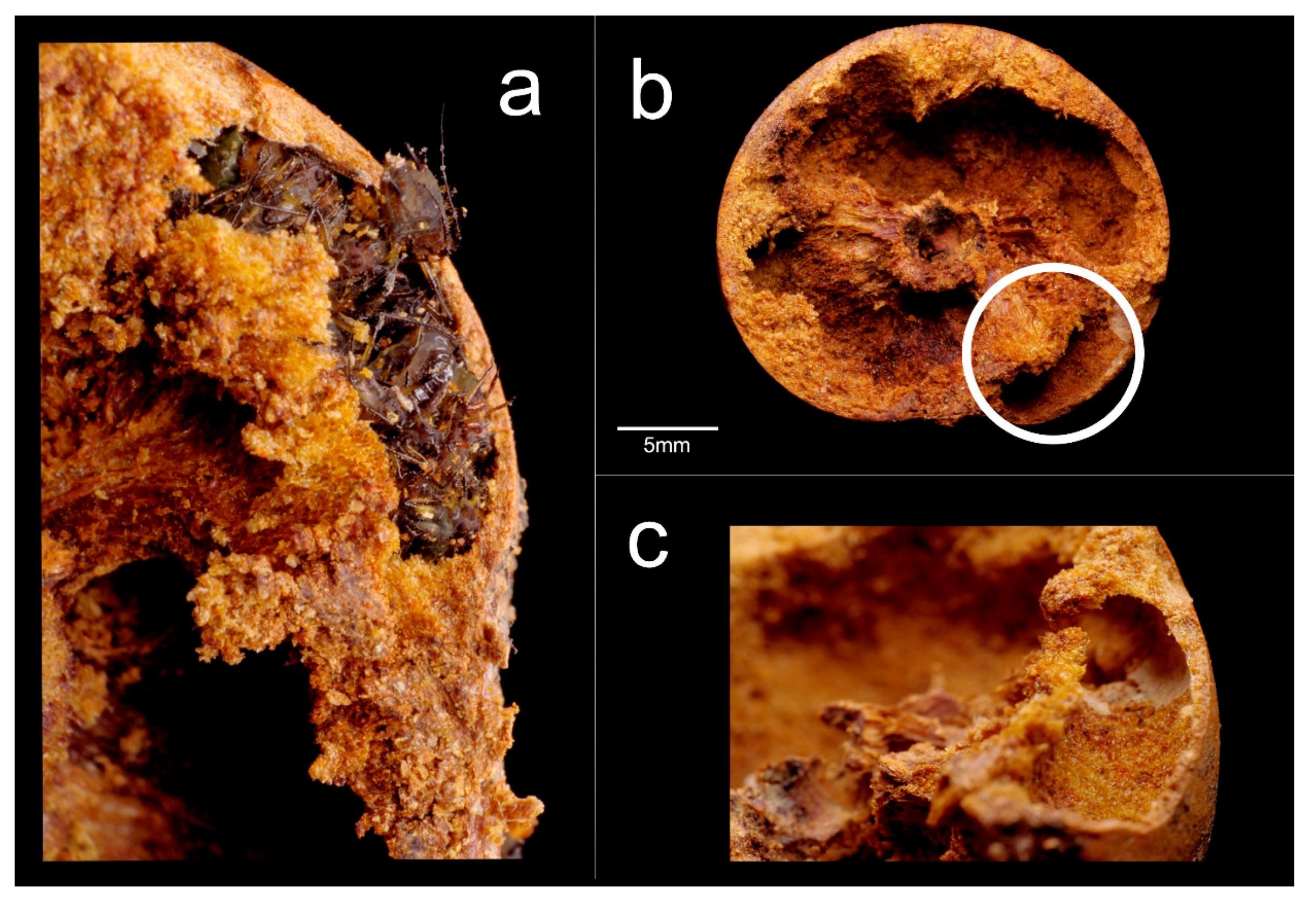
- Document which ant species colonized galls, what kind of colony composition characterized the gall-colonizing colonies (whether queens, workers or immature stages are present), and whether the galls’ height was a significant factor for colonization by different ant species.
- Document and quantify the presence of aphid species, suggested by some preliminary observation, and its relation to ant presence.
- Document how the presence of gall-colonizing ants or aphids influenced the galls’ internal architecture.
2. Materials and Methods
2.1. Study Area and Gall Selection
2.2. Survey
2.3. Field Experiment
2.4. Determination of Aphid Species by DNA Barcoding
2.5. Identification of Ant Species
2.6. Inner Architecture: Excavation Volume and 2D Analysis
2.7. Statistical Analyses
2.7.1. Survey
2.7.2. Field Experiment
2.7.3. Excavation Volume Analysis
2.7.4. Two-Dimensional Analysis (Nest Architecture)
3. Results
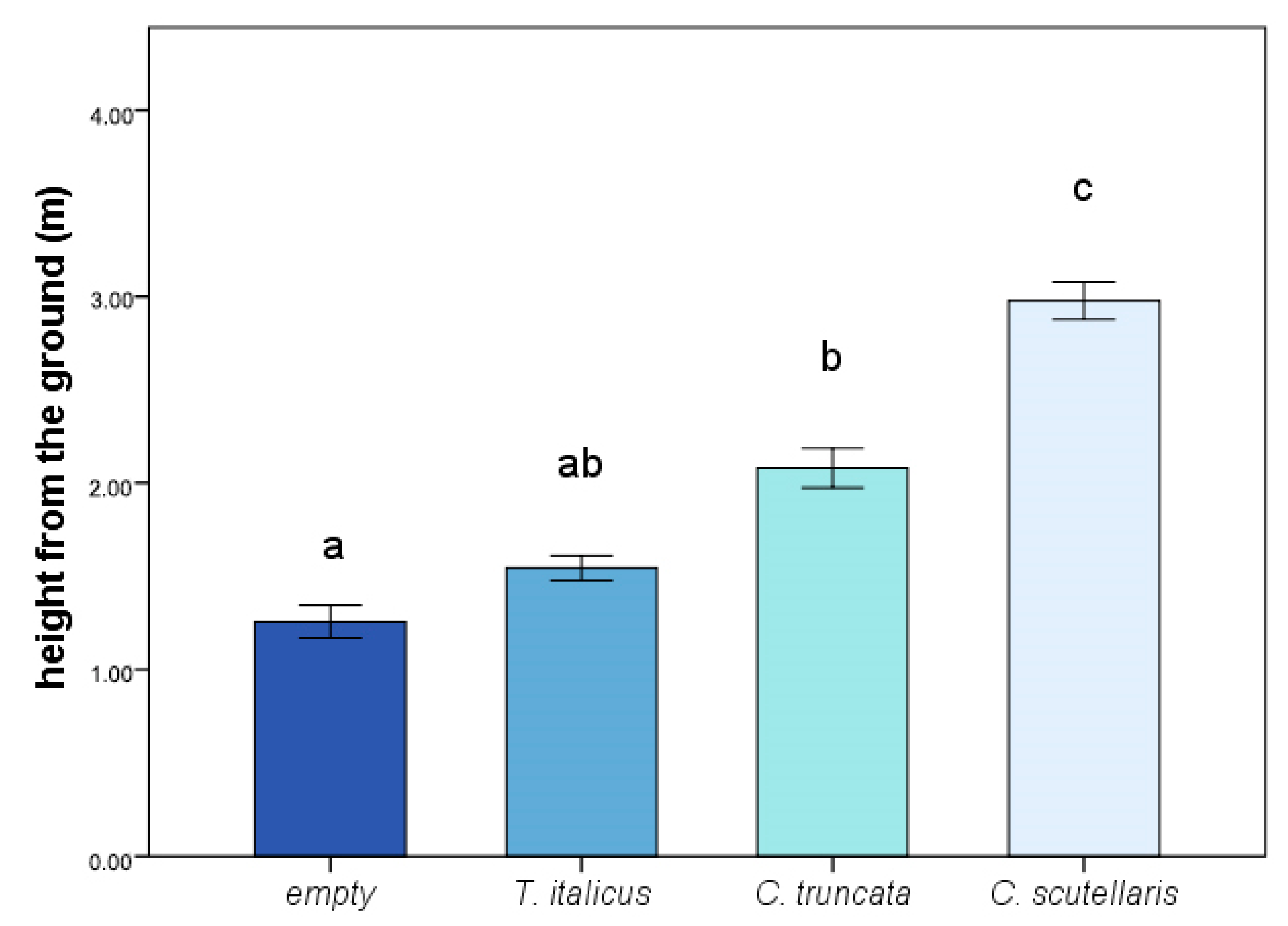
3.1. Survey
3.2. Field Experiment
3.3. Determination of Aphid Species by DNA Barcoding
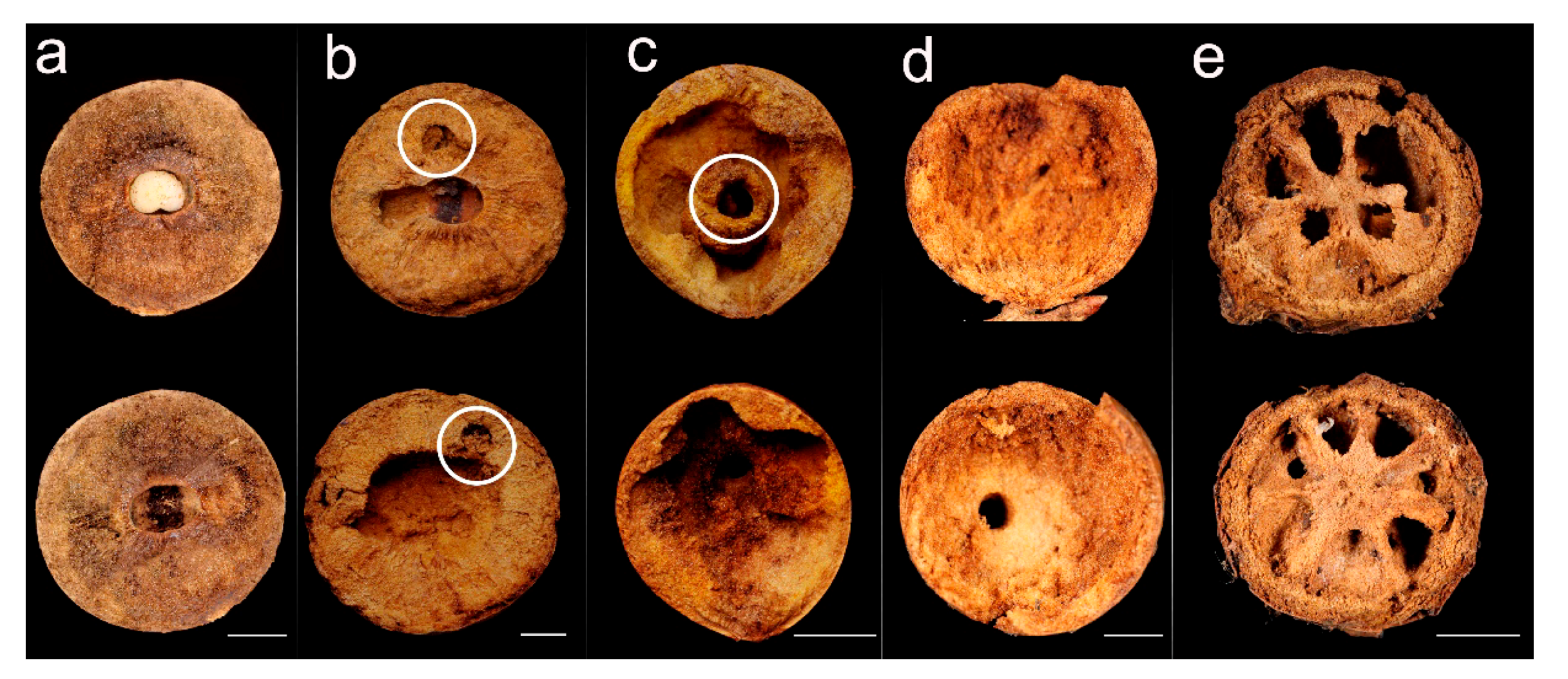
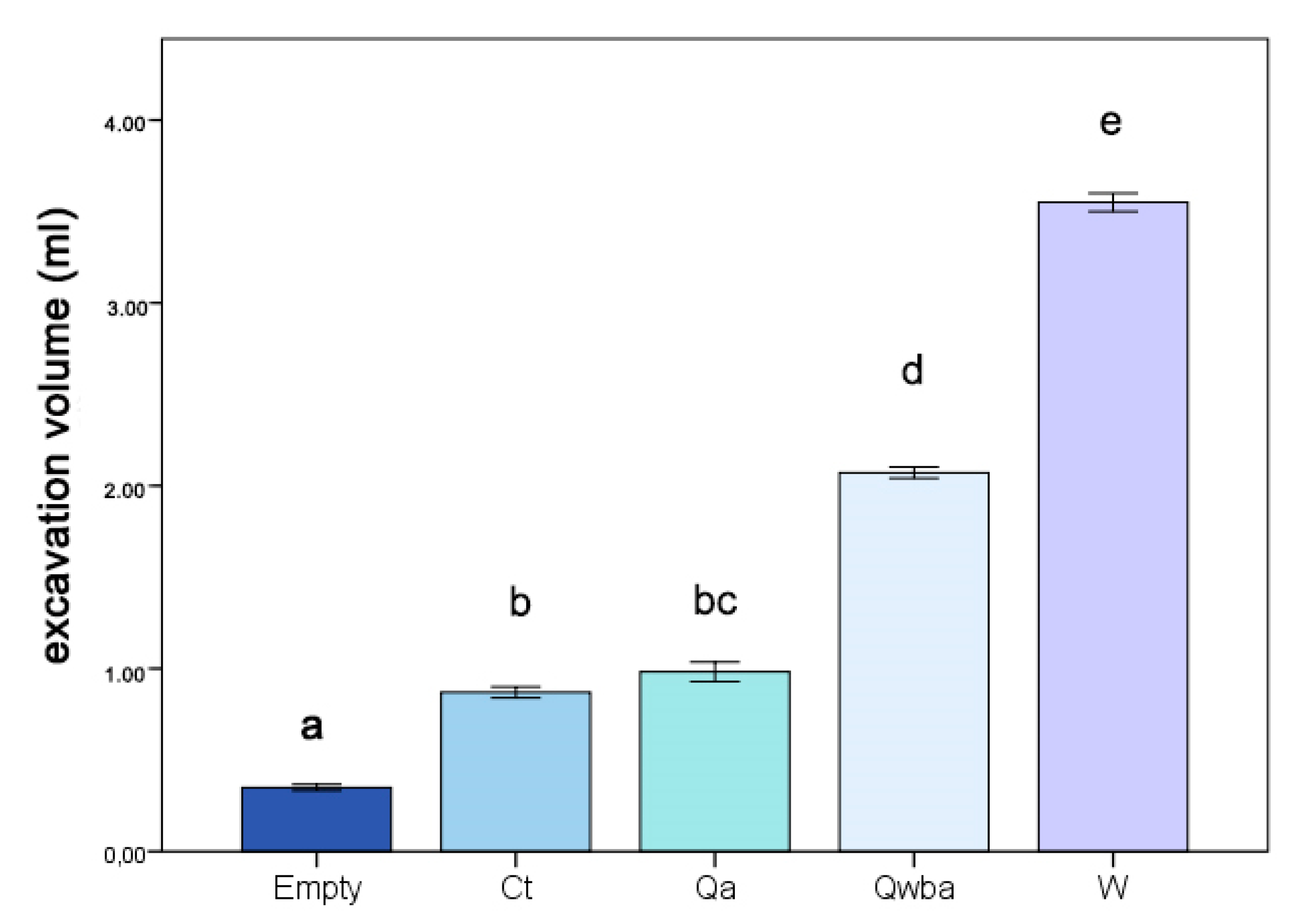
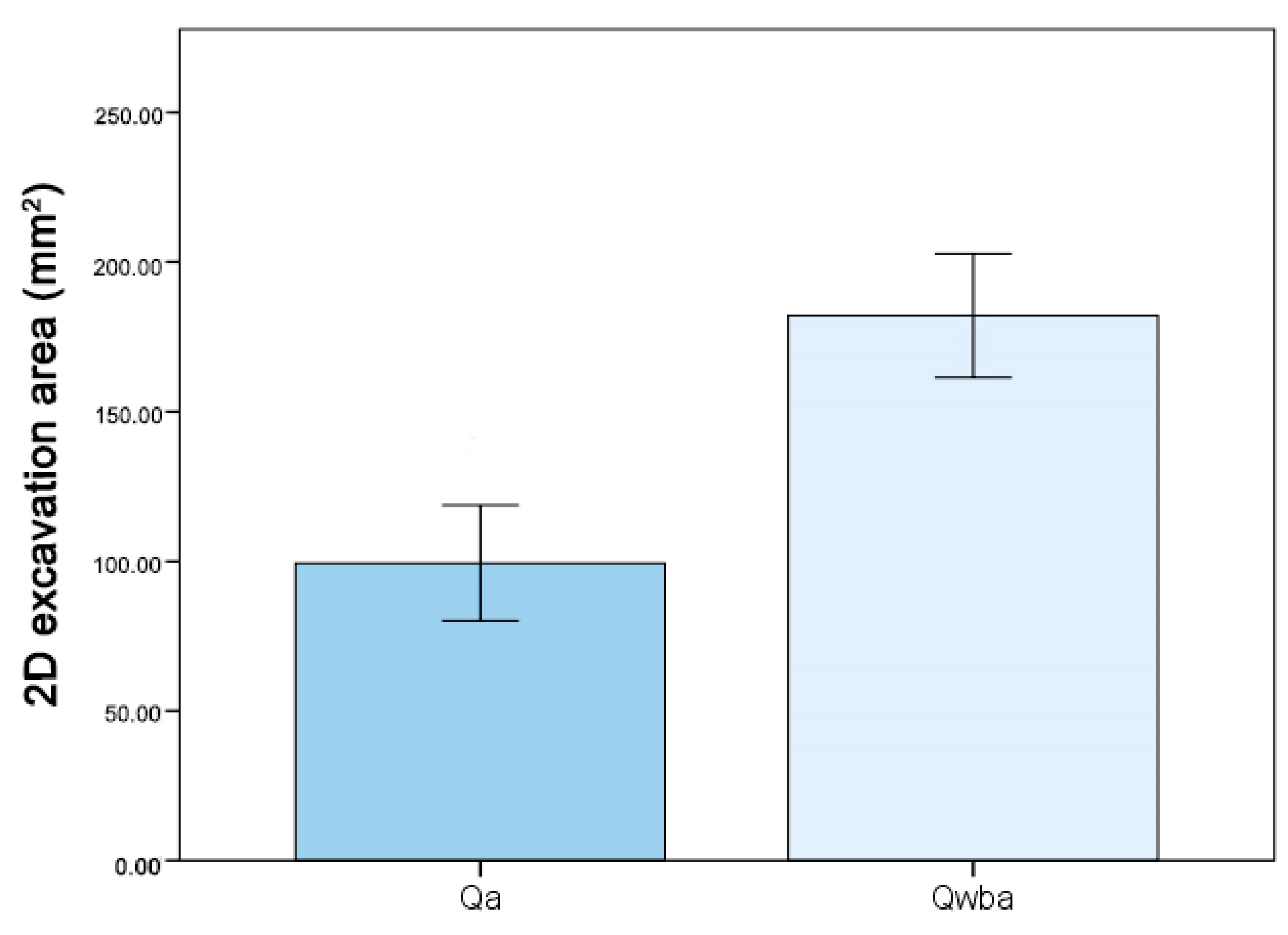
3.4. Inner Architecture: Excavation Volume and 2D Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Redfern, M.; Shirley, P.; Bloxham, M. British Plant Galls: Identification of Galls on Plants and Fungi; FSC Publications: Shrewsbury, UK, 2002. [Google Scholar]
- Price, W.P. Adaptive radiation of gall-inducing insects. Basic Appl. Ecol. 2005, 6, 413–421. [Google Scholar] [CrossRef]
- Harris, M.O.; Pitzschke, A. Plants make galls to accommodate foreigners: Some are friends, most are foes. New Phytol. 2020, 225, 1852–1872. [Google Scholar] [CrossRef] [PubMed]
- Giannetti, D.; Castracani, C.; Spotti, F.A.; Mori, A.; Grasso, D.A. Gall-Colonizing Ants and Their Role as Plant Defenders: From ‘Bad Job’ to ‘Useful Service’. Insects 2019, 10, 392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibb, H.; Dunn, R.R.; Sanders, N.J.; Grossman, B.F.; Photakis, M.; Abril, S.; Agosti, D.; Andersen, A.N.; Angulo, E.; Armbrecht, I.; et al. A global database of ant species abundances. Ecology 2017, 98, 883–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lach, L.; Parr, C.L.; Abbot, K.L. Ant Ecology; Oxford University Press: Oxford, UK, 2010. [Google Scholar]
- Visicchio, R.; Mori, A.; Grasso, D.A.; Castracani, C.; Le Moli, F. Glandular sources of recruitment, trail, and propaganda semiochemicals in the slave-making ant Polyergus Rufescens. Ethol. Ecol. Evol. 2001, 13, 361–372. [Google Scholar] [CrossRef]
- Grasso, D.A.; Mori, A.; Le Moli, F. Chemical communication during foraging in the harvesting ant Messor capitatus (Hymenoptera, Formicidae). Insectes Soc. 1998, 45, 85–96. [Google Scholar] [CrossRef]
- Oliver, T.H.; Leather, S.R.; Cook, J.M. Macroevolutionary patterns in the origin of mutualisms involving ants. J. Evol. Biol. 2008, 21, 1597–1608. [Google Scholar] [CrossRef] [Green Version]
- Nepi, M.; Grasso, D.A.; Mancuso, S. Nectar in plant-insect mutualistic relationships: From food reward to partner manipulation. Front. Plant Sci. 2018, 9, 1063. [Google Scholar] [CrossRef]
- Stadler, B.; Dixon, A.F. Mutualism: Ants and Their Insect Partners; Cambridge University Press: Cambridge, UK, 2008. [Google Scholar]
- Hölldobler, B.; Wilson, E.O. The Ants; Harvard University Press: Cambridge, UK, 1990. [Google Scholar]
- Delabie, J.H. Trophobiosis between Formicidae and Hemiptera (Sternorrhyncha and Auchenorrhyncha): An overview. Neotrop. Entomol. 2001, 30, 501–516. [Google Scholar] [CrossRef] [Green Version]
- Mandrioli, M.; Bisanti, M.; Grasso, D.A.; Manicardi, G.C. Role of ant-tending in modulating the presence of symbiotic bacteria against parasitoids in aphids. Trends Entomol. 2016, 12, 63–71. [Google Scholar]
- Kiers, E.T.; West, S.A. Evolving new organisms via symbiosis. Science 2015, 348, 392–394. [Google Scholar] [CrossRef] [PubMed]
- Douglas, A.E. The Symbiotic Habit; Princeton University Press: Princeton, NJ, USA, 2010. [Google Scholar]
- Gilbert, S.F. Ecological developmental biology: Environmental signals for normal animal development. Evol. Dev. 2012, 14, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Grasso, D.A.; Pandolfi, C.; Bazihizina, N.; Nocentini, D.; Nepi, M.; Mancuso, S. Extrafloral-nectar-based partner manipulation in plant–ant relationships. AoB Plants 2015, 7, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stadler, B.; Dixon, A.F. Ecology and evolution of aphid-ant interactions. Annu. Rev. Ecol. Evol. Syst. 2005, 36, 345–372. [Google Scholar] [CrossRef]
- Shingleton, A.W.; Stern, D.L. Molecular phylogenetic evidence for multiple gains or losses of ant mutualism within the aphid genus Chaitophorus. Mol. Phylogenet. Evol. 2003, 26, 26–35. [Google Scholar] [CrossRef]
- Nováková, E.; Hypša, V.; Klein, J.; Foottit, R.G.; von Dohlen, C.D.; Moran, N.A. Reconstructing the phylogeny of aphids (Hemiptera: Aphididae) using DNA of the obligate symbiont Buchnera aphidicola. Mol. Phylogenet. Evol. 2013, 68, 42–54. [Google Scholar] [CrossRef] [PubMed]
- Billick, I.; Hammer, S.; Reithel, J.S.; Abbot, P. Ant-aphid interactions: Are ants friends, enemies, or both? Ann. Entomol. Soc. Am. 2007, 100, 887–892. [Google Scholar] [CrossRef]
- Dixon, A.F.G. Aphid Ecology; Chapman and Hall: London, UK, 1998. [Google Scholar]
- Matsuura, K.; Yashiro, T. Aphid egg protection by ants: A novel aspect of the mutualism between the tree-feeding aphid Stomaphis hirukawai and its attendant ant Lasius productus. Naturwissenschaften 2006, 93, 506–510. [Google Scholar] [CrossRef]
- Ness, J.; Mooney, K.; Lach, L. Ants as mutualists. In Ant Ecology; Lach, L., Parr, C.L., Abbott, K.L., Eds.; Oxford University Press: New York, NY, USA, 2010; pp. 97–114. [Google Scholar]
- Ivens, A.B.; Kronauer, D.J.; Pen, I.; Weissing, F.J.; Boomsma, J.J. Ants farm subterranean aphids mostly in single clone groups-an example of prudent husbandry for carbohydrates and proteins? BMC Evol. Biol. 2012, 12, 106. [Google Scholar] [CrossRef] [Green Version]
- Styrsky, J.D.; Eubanks, M.D. Ecological consequences of interactions between ants and honeydew-producing insects. Proc. Biol. Sci. 2007, 274, 151–164. [Google Scholar] [CrossRef] [Green Version]
- Shingleton, A.W.; Stern, D.L.; Foster, W.A. The origin of a mutualism: A morphological trait promoting the evolution of ant-aphid mutualisms. Evolution 2005, 59, 921–926. [Google Scholar] [CrossRef] [PubMed]
- Kindlmann, P.; Hullé, M.; Stadler, B. Timing of dispersal: Effect of ants on aphids. Oecologia 2007, 152, 625–631. [Google Scholar] [CrossRef] [PubMed]
- Strathdee, A.T.; Bale, J.S.; Strathdee, F.C.; Block, W.C.; Coulson, S.J.; Webb, N.R.; Hodkinson, I.D. Climatic severity and the response to temperature elevation of Arctic aphids. Glob. Chang. Biol. 1995, 1, 23–28. [Google Scholar] [CrossRef]
- Leather, S.R. Egg survival in the bird cherry-oat aphid, Rhopalosiphum padi. Entomol. Exp. Appl. 1980, 27, 96–97. [Google Scholar] [CrossRef]
- Leather, S.R.; Dixon, A.F.G. The effect of cereal growth stage and feeding site on the reproductive activity of the bird-cherry aphid, Rhopalosiphum padi. Ann. Appl. Biol. 1981, 97, 135–141. [Google Scholar] [CrossRef]
- Wade, F.A.; Leather, S.R. Overwintering of the sycamore aphid, Drepanosiphum platanoidis. Entomol. Exp. Appl. 2002, 104, 241–253. [Google Scholar] [CrossRef]
- Hales, D.F.; Tomiuk, J.; Wöhrmann, K.; Sunnucks, P. Evolutionary and genetic aspects of aphid biology: A review. Eur. J. Entomol. 1997, 94, 1–55. [Google Scholar]
- Griffiths, E.; Wratten, S.D. Intra-and inter-specific differences in cereal aphid low-temperature tolerance. Entomol. Exp. Appl. 1979, 26, 161–167. [Google Scholar] [CrossRef]
- Leather, S.R. Aspects of aphid overwintering (Homoptera: Aphidinea: Aphididae). Entomol. Gen. 1992, 17, 101–113. [Google Scholar] [CrossRef]
- Schifani, E.; Castracani, C.; Giannetti, D.; Spotti, F.A.; Reggiani, R.; Leonardi, S.; Mori, A.; Grasso, D.A. New Tools for Conservation Biological Control: Testing Ant-Attracting Artificial Nectaries to Employ Ants as Plant Defenders. Insects 2020, 11, 129. [Google Scholar] [CrossRef] [Green Version]
- Seifert, B. The Ants of Central and Northern Europe; Lutra Verlags-und Vertriebsgesellschaft: Tauer, Germany, 2018. [Google Scholar]
- Foottit, R.G.; Maw, H.V.; Von Dohlen, C.D.; Hebert, P.D.N. Species identification of aphids (Insecta: Hemiptera: Aphididae) through DNA barcodes. Mol. Ecol. Resour. 2008, 8, 1189–1201. [Google Scholar] [CrossRef] [PubMed]
- Consani, M.; Zangheri, P. Fauna di Romagna. Imenotteri—Formicidi. Memorie Della Società Entomologica Italiana 1952, 31, 38–48. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. arXiv 2014, arXiv:1406.5823. [Google Scholar]
- Russell, V.L. Least-Squares Means: The R Package lsmeans. J. Stat. Softw. 2016, 69, 1–33. [Google Scholar]
- Offenberg, J.; Damgaard, C. Ants suppressing plant pathogens: A review. Oikos 2019, 128, 1691–1703. [Google Scholar] [CrossRef]
- Cammell, M.E.; Way, M.J.; Paiva, M.R. Diversity and structure of ant communities associated with oak, pine, eucalyptus and arable habitats in Portugal. Insectes Soc. 1996, 43, 37–46. [Google Scholar] [CrossRef]
- Santini, G.; Tucci, L.; Ottonetti, L.; Frizzi, F. Competition trade-offs in the organisation of a Mediterranean ant assemblage. Ecol. Entomol. 2007, 32, 319–326. [Google Scholar] [CrossRef]
- Solida, L.; Grasso, D.A.; Celant, A.; Fanfani, A.; Mori, A.; Le Moli, F. Foraging activity in two species of Messor harvester ants: Preliminary data on size-matching and diet breadth. Redia 2007, 90, 71–73. [Google Scholar]
- Solida, L.; Scalisi, M.; Fanfani, A.; Mori, A.; Grasso, D.A. Interspecific space partitioning during the foraging activity of two syntopic species of Messor harvester ants. J. Biol. Res. 2010, 13, 3–12. [Google Scholar]
- Santini, G.; Ramsay, M.P.; Tucci, L.; Ottonetti, L.; Frizzi, F. Spatial patterns of the ant Crematogaster scutellaris in a model ecosystem. Ecol. Entomol. 2011, 36, 625–634. [Google Scholar] [CrossRef]
- Maschwitz, U.; Hänel, H. The migrating herdsman Dolichoderus (Diabolus) cuspidatus: An ant with a novel mode of life. Behav. Ecol. Sociobiol. 1985, 17, 171–184. [Google Scholar]
- Collins, C.M.; Leather, S.R. Ant-mediated dispersal of the black willow aphid. Ecol. Entomol. 2002, 27, 238–241. [Google Scholar] [CrossRef]
- Jaśkiewicz, B.; Cichocka, E. Aphids on European Walnut (Juglans regia L.) in the urban conditions of Lublin. Aphids Other Hemipterous Insects 2004, 10, 35–46. [Google Scholar]
- Wani, S.A.; Tariq, A.S. Competition and niche-partitioning in twos of walnut. J. Sci. Res. Rev. 2014, 3, 120–125. [Google Scholar]
- Karczmarz, K. Dynamics of population and bionomics of Panaphis juglandis (Goeze, 1778) (Homoptera, Phyllaphididae) on common walnut (Juglans regia L.) in Lublin’s parks and gardens. Acta Sci. Pol. Hortorum Cultus 2012, 11, 53–70. [Google Scholar]
- Fremlin, M. The large walnut aphid (Panaphis juglandis Goeze)—A few observations. Nat. North East Essex 2016, 68–76. [Google Scholar]
- Stalažs, A.; Baka, I.; Svilāns, A. New species of three monophagous arthropods (Acari & Hemiptera) associated with common walnut (Juglans regia) in Latvia. Environ. Exp. Biol. 2020, 18, 1–6. [Google Scholar]
- Kök, Ş.; Aktaç, N.; Özdemir, I.; Kasap, I. The new association records on ants (Hymenoptera: Formicidae) and aphids (Hemiptera: Aphididae) in the Central Province of Çanakkale. Bitki Koruma Bülteni 2018, 58, 47–53. [Google Scholar]
- Czechowski, W.; Trigos-Peral, G.; Maák, I.; Vepsäläinen, K. Alate gyne of the ant Dolichoderus quadripunctatus (L.) (Hymenoptera, Formicidae) follows foraging trail to aphids. J. Hymenopt. Res. 2019, 71, 241–248. [Google Scholar] [CrossRef]
- Jakiewicz, B. Zdobniczka orzechowa (Panaphis juglandis Goetze) i zdobniczka podli-ciowa (Chromaphis juglandicola Kalt.)—Mszyce obniajce dekoracyjnoorzecha włoskie-go. Ochrona Rolin 2003, 10, 17–18. [Google Scholar]
- Wani, S.A.; Ahmad, S.T. Phenomenon of parthenogenesis, viviparity and endosymbiosis in aphids—A review. J. Bio. Innov. 2014, 3, 206–215. [Google Scholar]
- Sakata, H. Density-dependent predation of the ant Lasius niger (Hymenoptera: Formicidae) on two attended aphids Lachnus tropicalis and Myzocallis kuricola (Homoptera: Aphididae). Res. Popul. Ecol. 1995, 37, 159–164. [Google Scholar] [CrossRef]
- Offenberg, J. Balancing between mutualism and exploitation: The symbiotic interaction between Lasius ants and aphids. Behav. Ecol. Sociobiol. 2001, 49, 304–310. [Google Scholar] [CrossRef]
- Rico-Gray, V.; Oliveira, P. The Ecology and Evolution of Ant-Plant Interactions; The University of Chicago Press: Chicago, IL, USA, 2007. [Google Scholar]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giannetti, D.; Mandrioli, M.; Schifani, E.; Castracani, C.; Spotti, F.A.; Mori, A.; Grasso, D.A. First Report on the Acrobat Ant Crematogaster scutellaris Storing Live Aphids in Its Oak-Gall Nests. Insects 2021, 12, 108. https://doi.org/10.3390/insects12020108
Giannetti D, Mandrioli M, Schifani E, Castracani C, Spotti FA, Mori A, Grasso DA. First Report on the Acrobat Ant Crematogaster scutellaris Storing Live Aphids in Its Oak-Gall Nests. Insects. 2021; 12(2):108. https://doi.org/10.3390/insects12020108
Chicago/Turabian StyleGiannetti, Daniele, Mauro Mandrioli, Enrico Schifani, Cristina Castracani, Fiorenza A. Spotti, Alessandra Mori, and Donato A. Grasso. 2021. "First Report on the Acrobat Ant Crematogaster scutellaris Storing Live Aphids in Its Oak-Gall Nests" Insects 12, no. 2: 108. https://doi.org/10.3390/insects12020108